
 Voici le deuxième volet de notre série consacrée à l’Oxalis à larges feuilles (Oxalis latifolia).
Voici le deuxième volet de notre série consacrée à l’Oxalis à larges feuilles (Oxalis latifolia).Dans ce billet, nous allons passer en revue les parties aériennes de la plante.
La hampe florale
Nous pourrions commencer par examiner la tige. Cela ne sera toutefois pas possible, car elle n’existe pas ! En effet, Oxalis latifolia est une plante acaule, ce qui signifie justement sans tige (du grec ancien kaulós, « tige »). L’axe qui porte les fleurs de notre plante n’est pas une tige, car il est dépourvu de feuilles. Les botanistes appellent cet axe une hampe florale ou un pédoncule.
Les pétioles soutenant les feuilles partent directement du sol, formant une touffe haute de 5 à 30 cm qui entoure le pédoncule un peu plus élevé.

Si nous prenons comme critère la présence ou l’absence d’une tige, nous pouvons diviser le genre Oxalis en deux : les espèces du premier groupe possèdent des tiges feuillées, comme l’Oxalis corniculé et l’Oxalis de Dillenius (Oxalis dillenii). Celles du second groupe n’ont pas de tiges mais des hampes florales ; outre l’Oxalis à larges feuilles, c’est aussi le cas de l’Oxalis petite oseille (Oxalis acetosella) et de l’Oxalis à larges feuilles.

L’Oxalis de Dillenius (Oxalis dillenii) possède des tiges feuillées.
L’inflorescence
L’inflorescence est une ombelle composée d’une dizaine de fleurs (de 5 à 15). Une ombelle car les pédicelles sont insérés au même niveau de la hampe florale (voir photo ci-dessous).
Cette ombelle se compose de trois parties : une fleur centrale (1), qui est la première à s’ouvrir, et deux groupes latéraux (2 et 3), dont les pédicelles sont soudés à la base.

1 : fleur centrale
2 et 3 : groupes latéraux
L’axe principal de l’inflorescence se terminant par une fleur (la fleur centrale), sa croissance est donc arrêtée : les botanistes parlent dans ce cas d’une inflorescence définie ou déterminée (par opposition à une inflorescence indéfinie ou indéterminée). Ils appellent cela une cyme (du grec ancien kuma, « ce qui est gonflé »). Et puisque tous les pédicelles partent d’un point commun, on peut préciser qu’il s’agit d’une cyme ombelliforme.

Les pédicelles ne sont dressés que pendant l’anthèse (période durant laquelle la fleur est fonctionnelle), et penchés avant et après celle-ci. Ils se redressent à nouveau durant l’ouverture du fruit 4.

Les pédicelles sont penchés avant et après l’anthèse.
Tournons-nous maintenant vers les sépales (photo suivante). Ils sont soudés à la base, bordés d’une marge membraneuse et portent à leur sommet deux excroissances orangées, allongées et presque parallèles.

1 : glandes orangées
2 : marge membraneuse
Ces excroissances (certains auteurs parlent à leur propos de glandes) sont des dépôts d’oxalate. Nous avons vu plus haut que d’autres espèces du genre en possèdent sur le revers des feuilles ; Oxalis latifolia en a également, mais ils se trouvent à la pointe de ses sépales 6.
La fleur d’Oxalis latifolia est pourvue normalement de 5 pétales et d’autant d’étamines. On observe toutefois fréquemment des individus disposant de six ou même sept pétales. La base de leur face interne est blanchâtre, veinée de vert.
Lors de la préfloraison, juste avant l’épanouissement de la fleur, les pétales sont disposés de manière spiralée.

Individu à 6 pétales.
La base de leur face interne est blanchâtre striée de vert.
Les pétales sont disposés en spirale lors de la préfloraison.
Les feuilles
 Nous avons déjà évoqué les feuilles d’Oxalis latifolia dans un billet précédent. Rappelons brièvement qu’elles sont trifoliées, chaque foliole ayant une forme presque triangulaire de 3 à 6 cm de côté mais dont le sommet est nettement émarginé (il présente une échancrure arrondie).
Nous avons déjà évoqué les feuilles d’Oxalis latifolia dans un billet précédent. Rappelons brièvement qu’elles sont trifoliées, chaque foliole ayant une forme presque triangulaire de 3 à 6 cm de côté mais dont le sommet est nettement émarginé (il présente une échancrure arrondie).
Leur face inférieure ne montre pas de granulation orangée, contrairement à celles de l’Oxalis articulé (O. articulata) ou de l’Oxalis chétif (O. debilis). Nous avions vu que ces marques correspondent à des dépôts d’oxalate.
Nyctinastie
Un aspect intéressant des Oxalis est la façon dont leurs feuilles se rabattent la nuit. Ce mouvement est appelée nyctinastie (du grec ancien núx « nuit » et nastós « compact »).
La nyctinastie peut également se traduire par la fermeture des pétales, mais nous nous concentrerons dans la suite de l’article sur le repliement des feuilles, la nyctinastie foliaire. Ce phénomène est fréquent chez les Oxalidacées et les Fabacées (la famille des Trèfles).

Les folioles de l’Oxalide droite (Oxalis stricta) repliées

 C’est ce que pensait Linné, dans son ouvrage intitulé justement Somnus Plantarum, « Le sommeil des plantes ».
C’est ce que pensait Linné, dans son ouvrage intitulé justement Somnus Plantarum, « Le sommeil des plantes ».
Pour répondre à cette question, il faut d’abord s’accorder sur ce que représente le sommeil.
Pour certains, les plantes ne peuvent pas dormir, car elles n’ont pas de système nerveux central, contrairement aux humains et aux autres animaux.
Le système nerveux, et plus précisément le cerveau, joue en effet un rôle essentiel dans les cycles veille-sommeil. C’est lui qui contrôle les changements dans notre respiration, notre pression sanguine et notre tonus musculaire 1.
D’autres, en revanche, insistent sur le fait que notre cycle veille-sommeil est simplement un processus biologique qui appartient aux rythmes circadiens (du latin circa diem, « presque un jour »). Ces rythmes suivent une périodicité d’environ 24 heures, basée sur la rotation de la Terre. Ils sont observés non seulement chez les animaux, mais aussi chez les plantes, les champignons, et même les bactéries.

Une fleur d’Oxalis acetosella à la tombée de la nuit
Les rythmes circadiens des plantes déterminent notamment le moment où elles déploient leurs fleurs, les referment, émettent des parfums, etc. La photosynthèse se règle également sur l’horloge circadienne.
Quoi qu’il en soit, le sommeil des plantes est très différent de celui des animaux. Comme le proclame le botaniste français Francis Hallé, les végétaux représentent pour nous humains le summum de l’altérité 2.
Les phytochromes
Le repliement des feuilles des Oxalis au crépuscule est donc un rythme circadien. Mais comment les plantes parviennent-elles à détecter que la nuit tombe et que le moment est venu de refermer leurs feuilles ? Ont-elles des yeux ?

En effet, les phytochromes sont des récepteurs (basés sur une protéine) que les plantes utilisent pour détecter la lumière rouge. Ils peuvent basculer entre deux formes, l’une nommée Pr (r : red), sensible au rouge, et l’autre Pfr (far red), percevant le rouge lointain. Ils ont été découverts dans de nombreux tissus : graines, feuilles, bourgeons, etc.

Phytochrome (© Jmol Development Team Wikimedia Commons)
Grâce à ces phytochromes, le végétal peut déceler une réduction de la quantité de lumière reçue, ainsi qu’une diminution du rapport entre le rouge et le rouge lointain, deux signaux l’informant de la fin de la journée.
 Nous vous avions déjà parlé des phytochromes dans le billet intitulé :
Nous vous avions déjà parlé des phytochromes dans le billet intitulé :
La lutte pour la lumière.
Le pulvinus
Les plantes détectent donc la tombée de la nuit grâce à leurs phytochromes. Il nous reste à expliquer le mouvement de leurs feuilles. C’est ici qu’intervient le pulvinus.
Pulvinus vient du latin pulvis « poussière, poudre », et par extension « coussin (contenant de la poussière) ».
Le pulvinus, appelé aussi coussinet foliaire, est un bourrelet situé à la base du pétiole ou du pétiolule (le petit pétiole de chaque foliole d’une feuille composée).

Les trois pulvinus à la base des folioles d’une feuille d’Oxalis latifolia.
Il est composé d’un centre où passent les vaisseaux conducteurs de la sève, entouré d’une couronne de cellules à la paroi cellulaire fine. Ce sont ces cellules qui sont à l’origine du mouvement des feuilles. Elles sont réparties en deux groupes disposés de part et d’autre du centre : l’extenseur et le fléchisseur.

Coupe transversale d’un pulvinus.
Les couleurs sont arbitraires.
La dilatation ou le rétrécissement de ces cellules provoquent le mouvement des pièces foliaires. Lorsque les cellules formant l’extenseur sont enflées (turgescentes), celles du fléchisseur sont flasques, et vice-versa.
Examinons ce qui se passe dans le cas d’une feuille d’Oxalis.
En a, les cellules de l’extenseur sont gonflées et soutiennent le pétiolule. Celles du fléchisseur sont contractées. La foliole se redresse.
En b, ce sont les cellules du fléchisseur qui sont turgescentes, tandis que celles de l’extenseur se sont vidées. Le pétiolule s’affaisse.


La direction des mouvements nyctinastiques varie en effet selon les espèces. Dans la liste de ces genres, dressée par Darwin en 1880, 37 ont leurs feuilles qui se redressent pour la nuit, tandis que 32 genres voient les leurs s’abaisser 3.
Quelques espèces présentent même des mouvements de torsion.

Les folioles de la Luzerne d’Arabie (Medicago arabica) se redressent durant la nuit.
Il nous reste à comprendre ce qui provoque ces changements rapides de turgescence des cellules.
Prenons une cellule de l’extenseur.
Une cellule végétale est protégée par une membrane, elle-même entourée d’une paroi. À l’intérieur, baignant dans le cytoplasme, se trouvent le noyau ainsi qu’une vacuole, un grand compartiment aux fonctions diverses.

Une cellule de l’extenseur en position de « sommeil »
La membrane est détachée de la paroi, et la vacuole n’occupe qu’un volume restreint. La cellule est flasque. C’est encore la nuit : l’extenseur est contracté, le pétiolule est courbé, la foliole est repliée vers le bas.

Le soleil se lève. Les phytochromes détectent une augmentation de la quantité de lumière reçue, ainsi qu’un accroissement du rapport entre le rouge et le rouge lointain. Ceci déclenche, par des processus non complètement élucidés, un afflux d’ions (principalement des ions potassium K+) dans les vacuoles des cellules de l’extenseur.
Cette plus grande concentration en ions entraîne à son tout une entrée d’eau par osmose.
L’osmose
L’osmose est le transfert d’un solvant (généralement de l’eau) à travers une membrane lorsqu’on se trouve en présence de deux solutions de concentration différente.
Sur le premier dessin ci-dessous, nous voyons deux compartiments remplis d’eau pure, et séparés par une membrane semi-perméable (perméable à l’eau mais pas aux solutés).
Sur le dessin suivant, nous versons un corps soluble (par exemple du sucre ou du sel) dans le premier compartiment. Nous observerons alors (dernier dessin) un flux d’eau provenant du milieu le moins concentré en solutés vers celui le plus concentré. C’est le phénomène de l’osmose.
Dans l’extenseur, les vacuoles se remplissent donc d’eau et occupent désormais la plus grande partie des cellules. Les membranes s’étendent et se collent aux parois, les cellules se dilatent.

L’eau provient du fléchisseur, qui subit au même moment le phénomène inverse. Ses cellules, perdant une bonne partie de leur liquide, se contractent.
Si le fléchisseur se rétrécit, l’extenseur lui se gonfle. Le pétiolule reprend une position horizontale ; la foliole se redresse, prête pour une journée de photosynthèse.

L’utilité de la nyctinastie foliaire
La liste de Darwin dont nous avons parlé ci-dessus a été mise à jour, et compte désormais 202 genres adeptes de la nyctinastie foliaire, appartenant à 38 familles. Les Fabacées sont nettement majoritaires avec 131 genres, suivies, de très loin, par les Astéracées (7 genres). Cet inventaire ne comprend pas que des dicotylédones ; on y trouve des monocotylédones, dont 6 genres de Poacées, et même des conifères du genre Abies et des fougères aquatiques (Marsilea) 5.
Une caractéristique qui saute aux yeux quand on parcourt la liste des espèces pratiquant la nyctinastie foliaire est sa grande diversité. On y trouve des plantes herbacées, des arbustes et des arbres ; des plantes adaptées aux environnements arides, aquatiques, ou intermédiaires ; des plantes des zones tropicales ou tempérées.

Les frondes de la fougère Marsilea minuta, vivant en Inde, ressemblent aux feuilles des Oxalis.
© Vengolis ; CC BY-SA 4.0
Quel est donc leur intérêt de replier et de déployer leurs feuilles à intervalles réguliers ? Il doit s’agir d’une adaptation utile, puisqu’elle a un coût. Tout mouvement, après tout, consomme de l’énergie.
Les scientifiques ne sont sûrs de rien, mais ils ont évidemment échafaudé des hypothèses.
Protéger les feuilles contre le froid ?
Darwin pensait que la nyctinastie foliaire permettait d’éviter les dommages causés par le gel, en réduisant l’exposition du limbe au ciel nocturne.
Cette explication pose cependant plusieurs problèmes. Citons le fait que les principaux partisans de ce mécanisme, les Fabacées, sont très présents sous les Tropiques, précisément là où des conditions très froides sont rarement rencontrées. Ensuite, les mouvements des feuilles s’atténuent au fur et à mesure que la température diminue et s’approche du seuil critique pour les tissus, contrairement donc à ce que l’on s’attendrait à observer si l’hypothèse de Darwin était exacte 7.
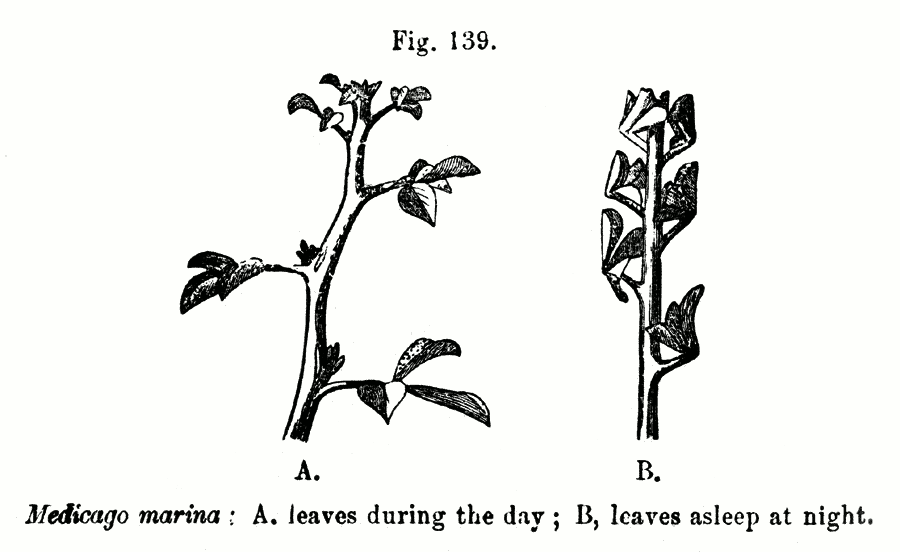
Mouvements foliaires chez Medicago marina, une Fabacée;
Illustration tirée du livre de Darwin « The Power of Movement in Plants »
Permettre l’écoulement de l’eau de pluie ?
Une autre raison qui est parfois avancée est qu’il s’agirait d’une adaptation facilitant l’écoulement de l’eau déposée par la pluie sur la surface des feuilles 9.
Le poids de l’eau retenue sur les feuilles pourrait en effet nécessiter un investissement plus important dans des tissus de soutien. Mais dans ce cas, les plantes présentant de la nyctinastie foliaire devraient dépenser moins de ressources dans le support des feuilles, et cela n’est pas avéré.

Certains chercheurs ont aussi suggéré qu’un film humide persistant sur le limbe pourrait constituer un habitat plus favorable pour les agents pathogènes. Le repliement des feuilles à la tombée de la nuit permettrait peut-être de diminuer les infections 10.
Se défendre contre les herbivores nocturnes ?
Selon d’autres spécialistes, la nyctinastie foliaire serait un moyen de défense contre les herbivores nocturnes 11.
Le pliage des feuilles rendrait leur accès plus difficile et diminuerait leur visibilité dans des conditions crépusculaires.
Cette explication pourrait élucider un mystère : les scientifiques recherchent toujours la raison de la popularité de la nyctinastie foliaire parmi les Fabacées. Le lien résiderait peut-être dans leur teneur élevée en azote, due à leur association symbiotique avec des bactéries du sol fixatrices de cet élément 12. Or, l’azote est un matériau essentiel des organismes vivants, et des feuilles riches en ce composant sont particulièrement attirantes pour les herbivores. Le repliement des feuilles constituerait donc pour les Fabacées une défense supplémentaire contre leurs prédateurs, en plus de toutes les armes chimiques dont elles disposent déjà.

Ceci explique qu’ils soient souvent utilisés comme engrais verts
Aider les ennemis des ennemis ?
Le but est le même que dans l’hypothèse précédente : se défendre contre les folivores. Mais on suppose ici que la nyctinastie foliaire agirait de manière indirecte, en facilitant la capture des herbivores par leurs prédateurs. Ce serait donc une relation tritrophique (entre trois niveaux de la chaîne alimentaire) : une plante, des phytophages et les consommateurs de ceux-ci.

Prenons d’abord le cas des rongeurs. Nombre d’entre eux utilisent le couvert végétal pour se déplacer d’un point de nourrissage à un autre, et passent plus de temps à manger lorsqu’ils sont protégés par la végétation. Si cet écran disparaît, leur capture devient naturellement plus aisée.
On peut aussi penser que la position verticale des feuilles réduit le bruit acoustique. Les chauves-souris et les rapaces nocturnes seraient donc mieux à même de détecter les sons et les échos émis par leurs proies sans les couches de feuillage qui amortissent et dispersent les signaux 13.

D’une manière analogue, la fermeture des feuilles permettrait aux odeurs émises par les insectes herbivores d’atteindre plus rapidement l’appareil olfactif de leurs prédateurs nocturnes 14.

Sources :
1 : Fink A.M. et al. ; Autonomic regulation during sleep and wakefulness: a review with implications for defining the pathophysiology of neurological disorders ; Clinical Autonomic Research ; volume 28 ; n° 6 ; 2018 ; pp. 509-518 ↑
2 : Francis Hallé « Une plante n’a ni queue ni tête, c’est pour cette altérité que je suis devenu botaniste » ; Libération ; 29 décembre 2016 ↑
3 : Darwin ; The Power of Movements in Plants ; John Murray ; London ; 1880 ↑
4 : Pierre Rivals ; Sur la vie et les problèmes de la destruction d’Oxalis latifolia Kunth ; Journal d’agriculture tropicale et de botanique appliquée ; volume 7 ; n° 9-10 ; septembre-octobre 1960 ; pp. 397-405 ↑
5 : Peter V. Minorsky ; The functions of foliar nyctinasty: a review and hypothesis ; Biological Reviews ; volume 94 ; n° 1 ; 2019 ; Appendice S1 ↑
6 : Guy L. Nesom ; Taxonomic notes on acaulescent Oxalis (Oxalidaceae) in the United States ; Phytologia ; décembre 2009 ; volume 91 ; n° 3 ; pp. 501-526 ↑
7 : Peter V. Minorsky ; The functions of foliar nyctinasty: a review and hypothesis ; Biological Reviews ; volume 94 ; n° 1 ; 2019 ↑
8 : Oberlander et al. ; A model of bulb evolution in the eudicot genus Oxalis (Oxalidaceae) ; Molecular Phylogenetics and Evolution ; volume 51 ; pp. 54–63 ; 2009 ↑
9 : Dean, J. M. & Smith, A. P. ; Behavioral and morphological adaptations of a tropical plant to high rainfall ; Biotropica ; Volume 10 ; pp. 152–154 ; 1978 ↑
10 : Gilbert & Reynolds ; Nocturnal fungi: Airborne spores in the canopy and understory of a tropical rain forest ; Biotropica ; Volume 37 ; pp. 462-464 ; 2005 ↑
11 : Grubb, P. J. & Jackson, R. V. ; The adaptive value of young leaves being tightly folded or rolled on monocotyledons in tropical lowland rain forest, an hypothesis in two parts ; Plant Ecology ; n° 192 ; pp. 317-327 ; 2007 ↑
12 : Grubb, P. J. & Jackson, R. V. ; The adaptive value of young leaves being tightly folded or rolled on monocotyledons in tropical lowland rain forest, an hypothesis in two parts ; Plant Ecology ; n° 192 ; pp. 317-327 ; 2007 ↑
13 : Arlettaz, R., Jones, G. & Racey, P. A. ; Effect of acoustic clutter on prey detection by bats ; Nature ; n° 414 ; pp. 742-745 ; 2001 ↑
14 : Aartsma, Y., Bianchi, F. J. J. A., van der Werf, W. & Poelman, E. H. ; Herbivore‐induced plant volatiles and tritrophic interactions across spatial scales ; New Phytologist ; n° 216 ; pp. 1054–1063 ; 2017 ↑

Nyctinastie, un terme que je découvre et magnifiquement développé dans cet article très intéressant. Merci de tout ce travail de compilation et de restitution.
J’aimeJ’aime
Merci pour cette re cherche impressionante et sa bibliographie ad hoc, passionant!
J’aimeJ’aime